




7月3日,生命科學(xué)學(xué)院楊仲南、朱駿教授團隊于《Plant Biotechnology Journal》發(fā)表了題為“Mutation of GMC oxidoreductase leads to thermosensitive genic male sterility in rice and Arabidopsis”的研究論文,該研究發(fā)現(xiàn)了一個新的水稻溫敏不育的遺傳位點,從細胞學(xué)層次解析了溫敏不育的機制,反映了水稻和擬南芥溫敏不育的共同性和差異性。

自上世紀80年代發(fā)現(xiàn)水稻光溫敏不育系以來,光溫敏不育系已經(jīng)廣泛應(yīng)用于水稻兩系雜交制種。經(jīng)過四十多年的努力,育種學(xué)家發(fā)現(xiàn)了十多個水稻光溫敏不育系。近年研究發(fā)現(xiàn),生產(chǎn)上應(yīng)用的水稻兩系光溫敏不育系中90%以上的品種都是利用現(xiàn)有的tms5位點。一些年份偶發(fā)的夏季低溫潮導(dǎo)致帶有該位點不育系育性部分恢復(fù),嚴重影響雜交制種的純度。因此農(nóng)業(yè)生產(chǎn)上迫切需要不育性更為穩(wěn)定的光溫敏不育系。課題組先前對模式植物擬南芥光溫敏不育機制開展了系統(tǒng)深入的研究,發(fā)現(xiàn)低溫下小孢子發(fā)育緩慢是育性恢復(fù)的共性機制(Zhu et al., 2020, Nature Plants 6:360-367)。光敏不育系育性恢復(fù)也是由短光照條件下小孢子發(fā)育緩慢所致,因此光照和溫度共同通過調(diào)控小孢子發(fā)育進程從而恢復(fù)育性(Zhang et al., 2020, Plant Physiology 184: 923-932)。課題組篩選到一系列光溫敏不育系的恢復(fù)子,揭示了低溫下緩慢發(fā)育降低小孢子對細胞壁保護作用的要求從而恢復(fù)育性的細胞學(xué)機制(Shi et al., 2021, Molecular Plant 14:2104-2114; Wang et al.,2021, Journal of Integrative Plant Biology 64: 717-730)。
課題組對擬南芥光溫敏不育機制深入解析的基礎(chǔ)上,發(fā)明了水稻光溫敏不育系的創(chuàng)制技術(shù),獲得了多個中花11(ZH11)為背景的光溫敏不育系。其中一個命名為ostms18的株系在高溫(HT)下(29 ℃)表現(xiàn)為完全不育,但在低溫(LT)下(23 ℃)育性正常,且其他農(nóng)藝性狀與野生型相比沒有明顯區(qū)別。在 2021 年上海地區(qū)偶發(fā)的夏季低溫潮的影響下,tms18 與同樣為中花 11 背景的 tms5突變體相比,其高溫不育性狀顯著好于tms5。通過將該位點引入不同背景的水稻品種中發(fā)現(xiàn),含有其純合突變位點的植株仍表現(xiàn)為雄性不育,因此該遺傳位點在兩系雜交水稻育種中具有較好的應(yīng)用潛力(圖1)。
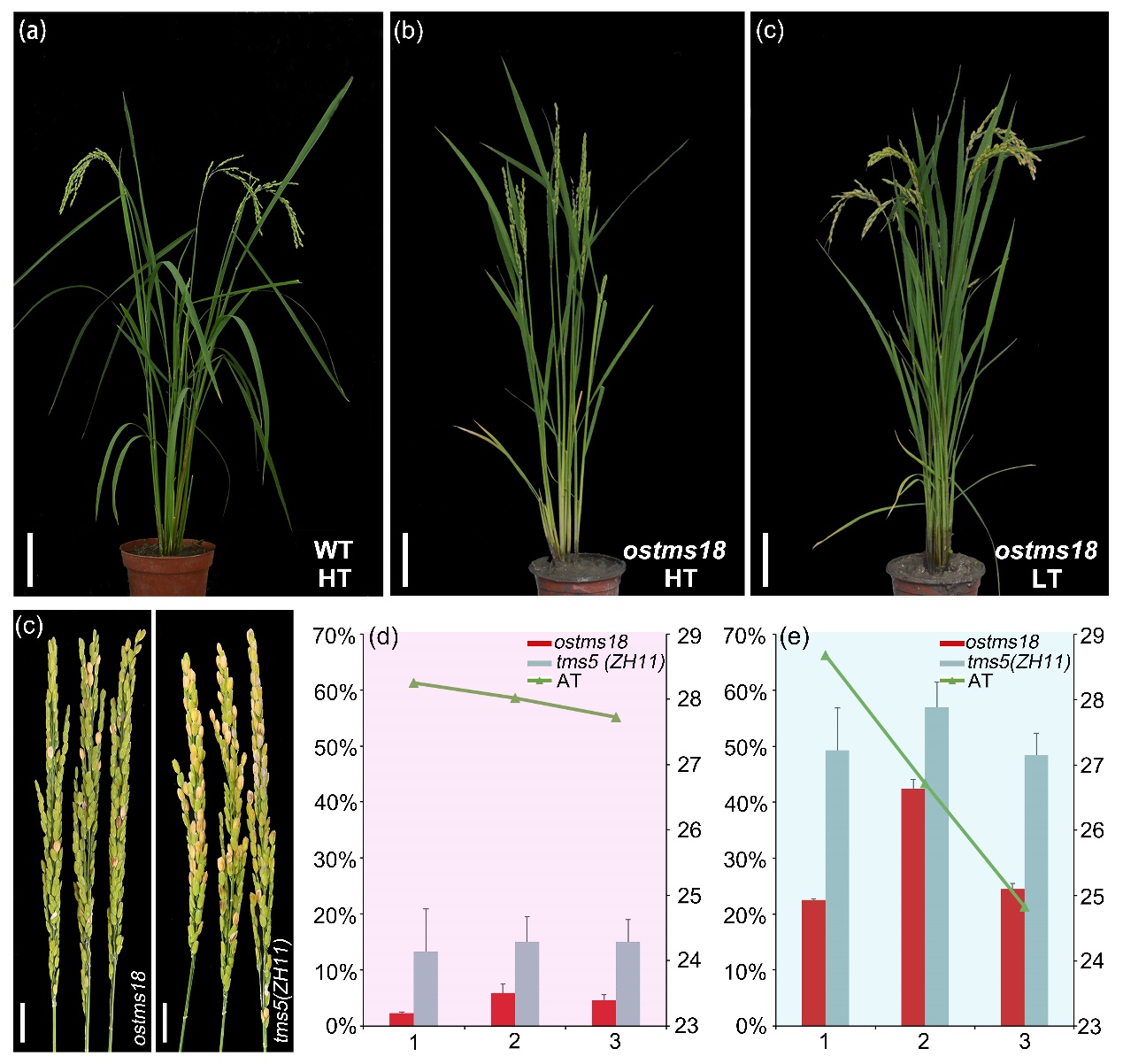
圖1. ostms18的溫敏不育性狀優(yōu)于同樣背景的tms5位點
OsTMS18編碼一個在花藥中高效表達的葡萄糖-甲醇-膽堿(GMC)氧化還原酶。先前其他課題組報道該基因強等位突變體np1為完全雄性不育。ostms18中該基因第二個外顯子一個氨基酸發(fā)生突變(Gly-Ser)導(dǎo)致光溫敏雄性不育。細胞學(xué)分析表明,ostms18 突變體在高溫下花粉外壁第二層的結(jié)構(gòu)發(fā)生異常,導(dǎo)致花粉破裂,但低溫下外壁第二層的結(jié)構(gòu)雖然變薄但較為完整。擬南芥中同源基因AtTMS18 的突變體在常溫下(24 ℃)育性正常,高溫條件下(28 ℃)育性顯著降低,同樣表現(xiàn)為溫敏不育性狀。此外,OsTMS18以及 AtTMS18基因分別受到了絨氈層花粉外壁形成關(guān)鍵轉(zhuǎn)錄因子 OsMS188 和 MS188 的直接調(diào)控,表明它們都參與到了花粉壁的形成過程中(圖2)。
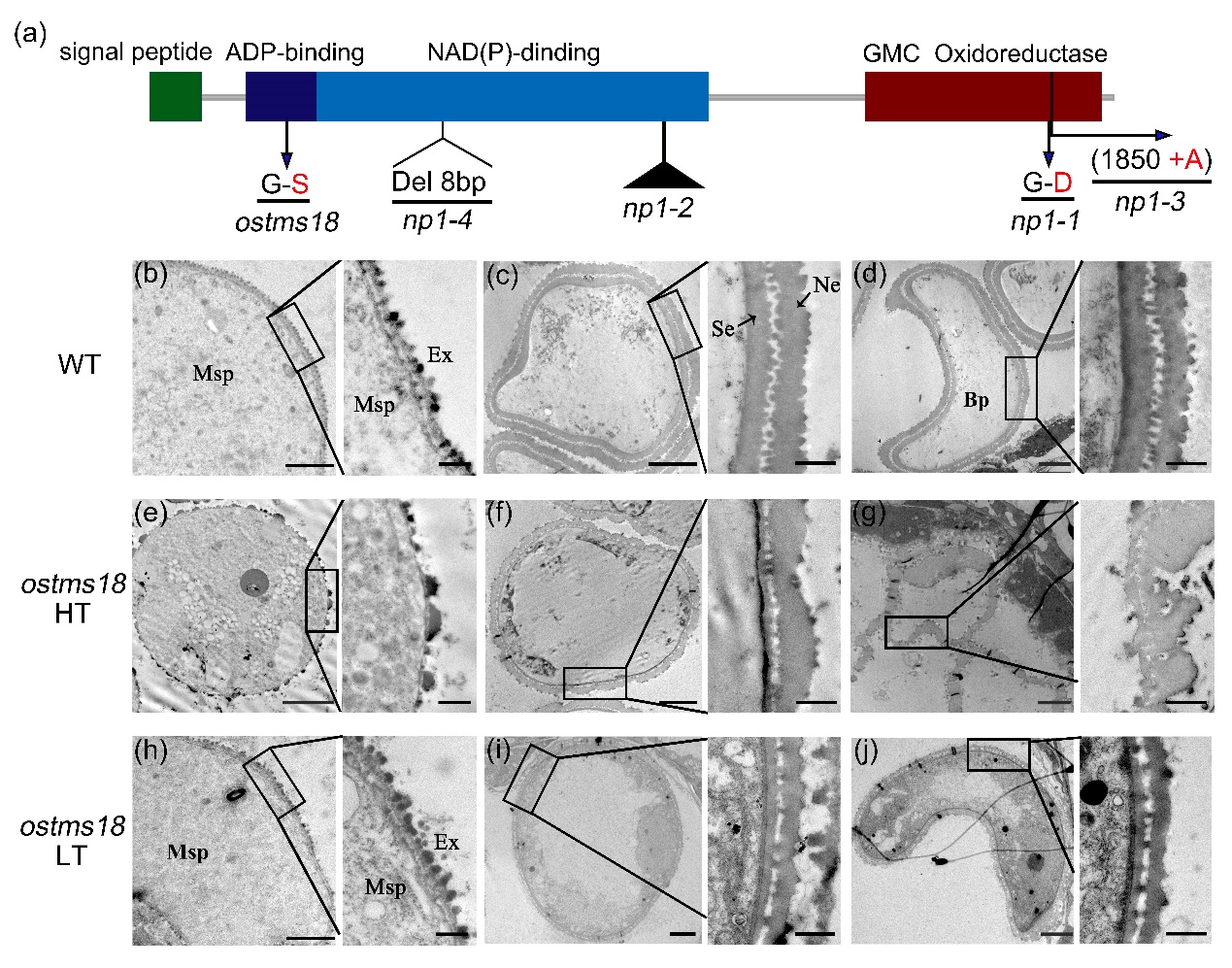
圖2. OsTMS18基因結(jié)構(gòu)及野生型和突變體在高低溫下的細胞學(xué)分析
基于以上研究結(jié)果,研究者提出了水稻光溫敏不育系ostms18育性恢復(fù)的細胞學(xué)機制。在小孢子發(fā)育成花粉過程中,需要進行細胞壁的轉(zhuǎn)換。四分體壁逐漸降解同時緊隨花粉外壁形成,隨后在花粉外壁和質(zhì)膜之間形成的花粉內(nèi)壁對成熟花粉粒起保護作用。ostms18中花粉外壁缺陷導(dǎo)致花粉破裂;低溫條件下的緩慢發(fā)育,有缺陷的花粉壁足以保護小孢子繼續(xù)形成花粉內(nèi)壁從而恢復(fù)育性。另外,該文章對于不同植物的光溫敏育性恢復(fù)的差異性進行了分析。水稻在夏季強日照和高溫條件下生長,花粉形成過程中需要更強的花粉壁保護。而擬南芥作為陰生模式植物,小孢子發(fā)育過程中對花粉壁保護的要求不高,相關(guān)基因的突變體育性更容易恢復(fù)。這些結(jié)果有助于對于植物光溫敏育性恢復(fù)機制的理解(圖3)。
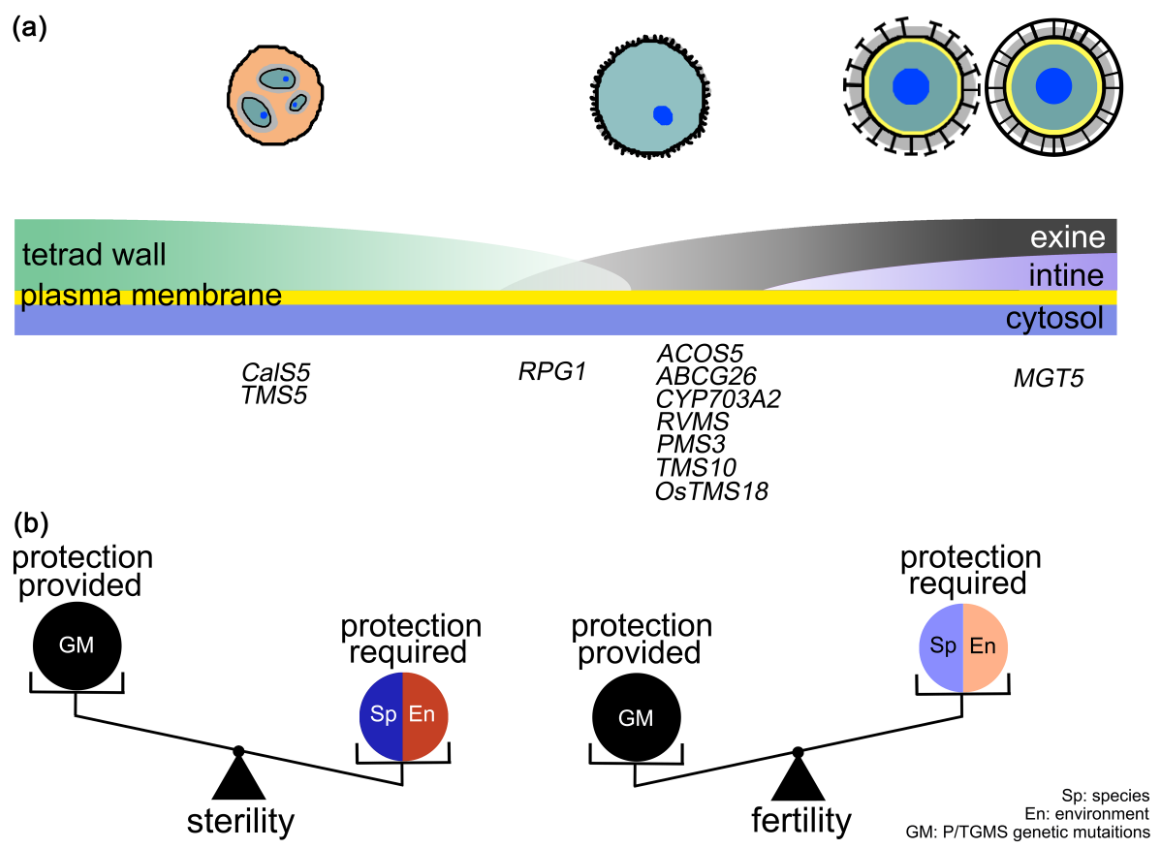
圖3. 植物光溫敏育性恢復(fù)的細胞水平機制
在該研究中,上海師范大學(xué)博士生張艷飛和李月靈博士為論文共同第一作者,楊仲南教授與朱駿教授為共同通訊作者,黃學(xué)輝教授也參與了部分研究工作。該研究得到了國家自然科學(xué)基金、上海市教委和科委等課題的資助。
文章鏈接:https://onlinelibrary.wiley.com/doi/10.1111/pbi.13886
(供稿、圖片:生命科學(xué)學(xué)院)





 徐匯校區(qū):上海市徐匯區(qū)桂林路100號
徐匯校區(qū):上海市徐匯區(qū)桂林路100號